基于楸树转录组数据开发了3,999对SSR引物并进行种间通用性验证(Wang等,2016;杨丹丹等,2018)。选择其中54对基于全长cDNA序列的基因开发的SSR标记,对108份梓属无性系进行基因分析;亲缘关系分析发现灰楸和滇楸在相同的分枝中表明她们为一个种,与最新版的《中国植物志》结论相同;群体结构分析将其分为7个亚群(图7)。
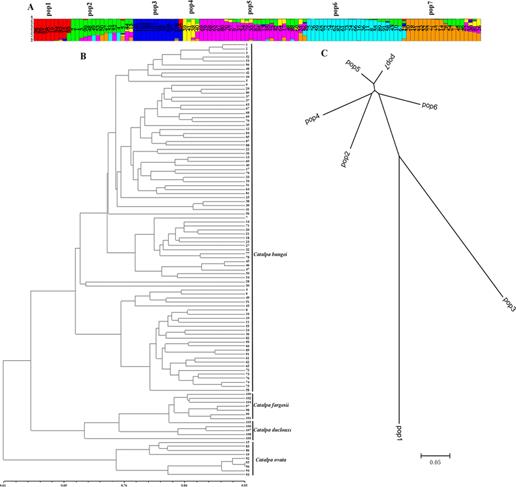
图7. 108份种质资源的遗传多样性分析(Wang等,2019,DNA and cell biology)
A群体结构,B种质资源亲缘关系,C亚群体亲缘关系
利用MLM模型分析不定根数、不定根长及不定根生根率表型数据和无性系的基因分型数据,检测到4个标记与不定根性状显著关联(p<0.01),解释表型贡献率的范围是10.13%~18.49%。在4个分子标记中,分子标记CBG010在两年都检测到与不定根数发生显著关联,贡献率分别为10.77%和18.49%。对CBG010的来源基因进行注释分析后发现其是一个WRKY转录因子基因,命名为CbNN1(Genban ID:AMP82887)(图8)
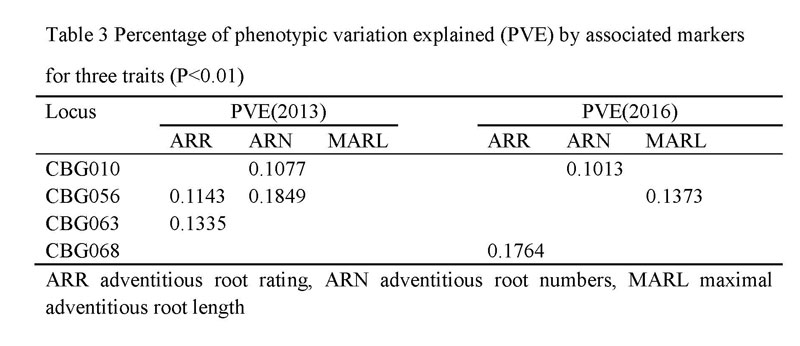
图8.关联分析检测的分子标记及贡献率(Wang等,2019,DNA and cell biology)
CbNN1基因的qRT-PCR分析发现在IBA诱导的楸树不定根发育的愈伤期及不定根伸长期该基因显著上调表达,组织表达分析发现在根、叶片、韧皮部、木质部都表达,根中表达量最高并且其表达量随着不定根生根率的增加而提高(图9,图10)。
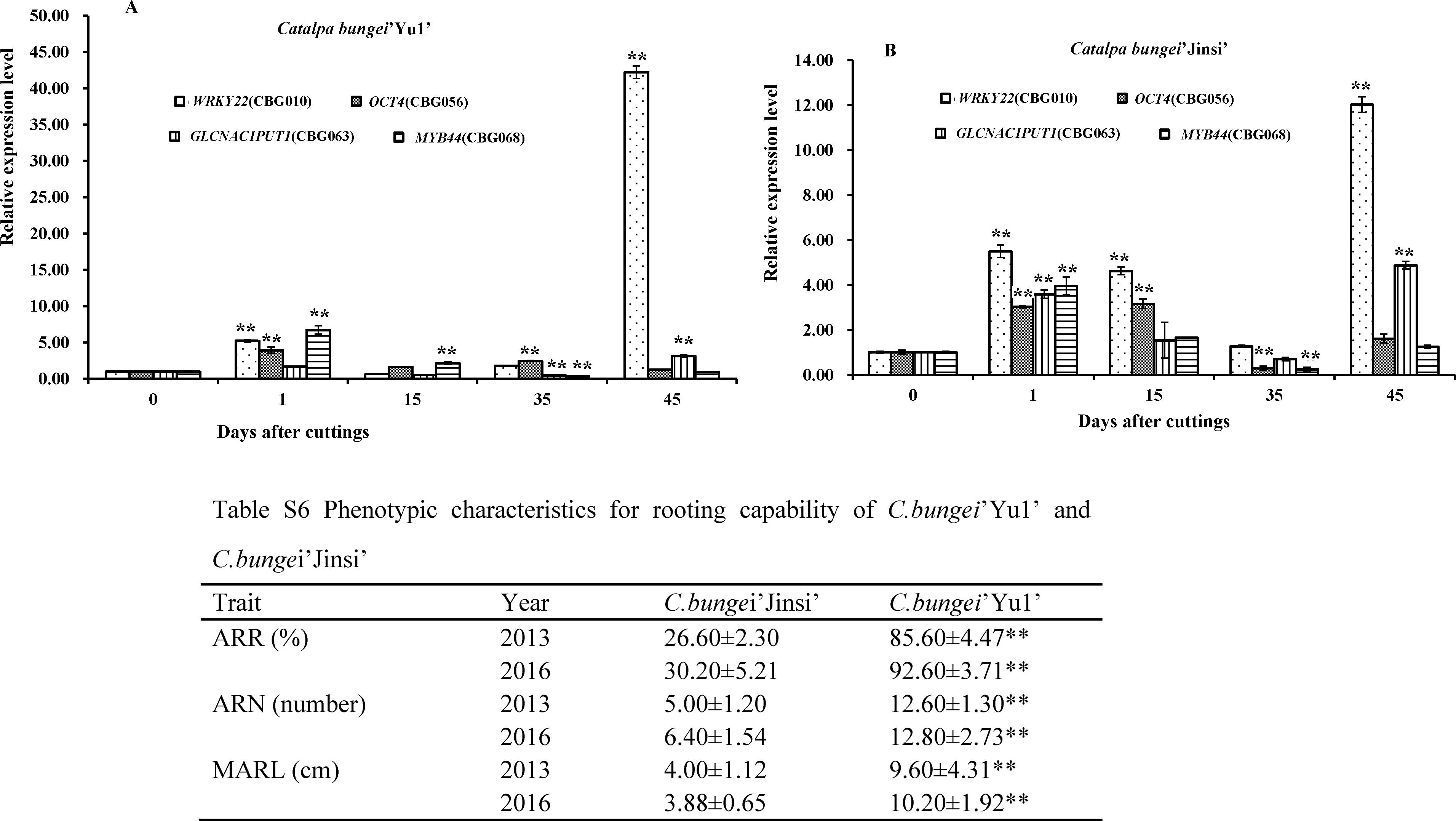
图9. CbNN1基因在不定根发育不同时期的表达分析(Wang等,2019,DNA and cell biology)
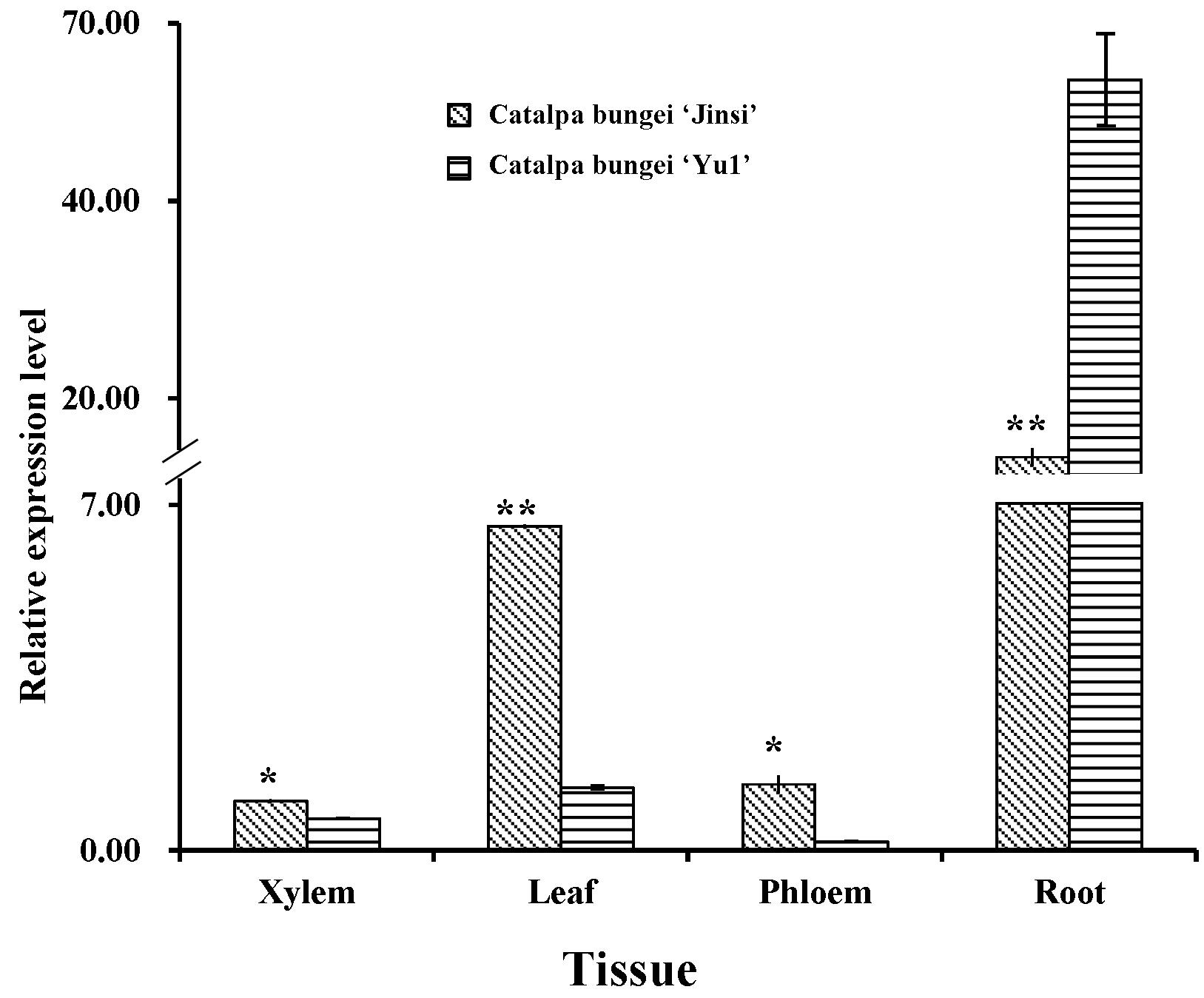
图10. CbNN1基因组织表达分析(Wang等,2019,DNA and cell biology)
根据‘豫楸1号’的survey序列和转录组序列,申请人设计引物,PCR扩增CbNN1基因的基因组序列。其基因组序列长度为1604 bp,ORF全长为1,056 bp,包括3个外显子,编码352个氨基酸。生物信息学分析发现其与拟南芥的AtWRKY22基因同源,具有一个WKRY结构域和一个CX5CX23HX1H锌指结构,核定位信号N端,序列为PRSKRRKNH,二级结构分析发现无规卷曲占到52.57%,其次是α螺旋(25.85%)(图11)。CbNN1基因的亚细胞定位分析发现其定位细胞核中,与预测的结果一致(图12)。
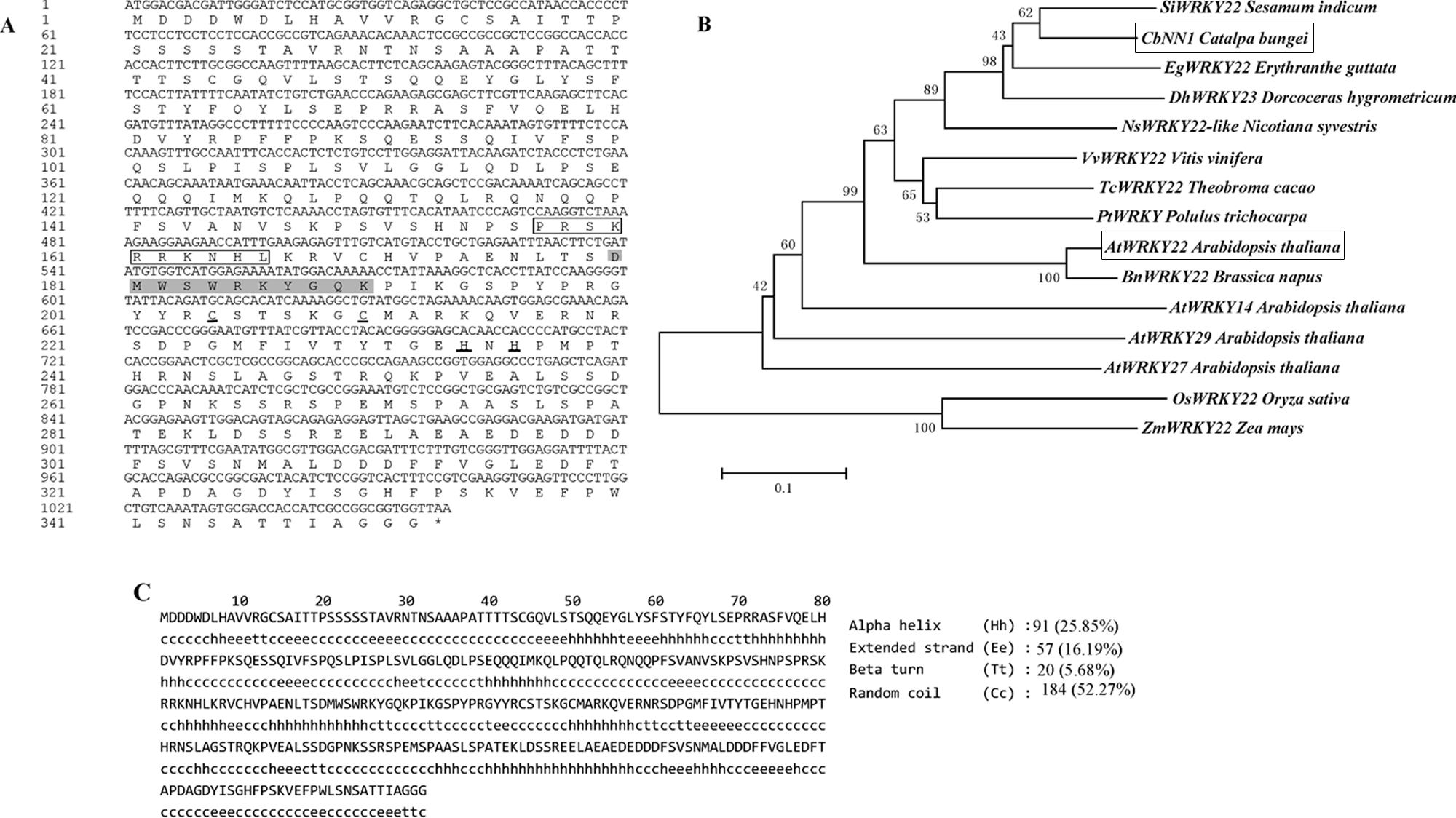
图11. CbNN1基因的生物信息学分析(Wang等,2019,DNA and cell biology)
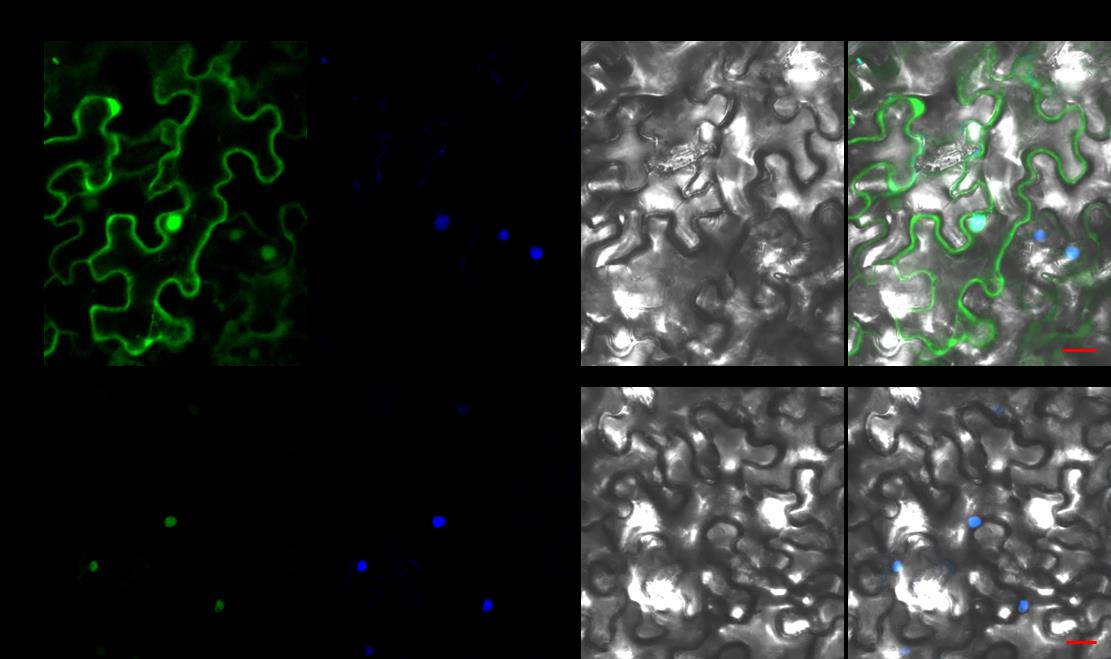
图12. CbNN1基因的亚细胞定位(bar=20μm)
根据‘豫楸1号’的survey序列,申请人克隆并分析了CbNN1基因上游1.65 kb启动子调控元件,分析发现G-box、TATA-box、ABRE、LTR、TCA等主要元件,表明CbNN1基因的表达可能主要受光照和激素诱导。

图12. CbNN1基因启动子顺式作用元件分析
利用已经构建的楸树遗传转化体系,申请人构建了CbNN1的过表达载体,经农杆菌介导的转化后获得转基因植株。转基因植株经过GUS染色和PCR扩增后获得CbNN1超表达转基因植株10株,选择其中的3株经组织培养增殖后(王鹏等,授权专利,2015),获得大量幼苗。幼苗在空白DKW培养基中培养10天后,移入生根培养基。在DKW基本培养基中,添加0 mg/L 的IBA或NAA时,转基因植株生根率可以达到45.44%-59.25%,空白对照则为0;添加0.1 mg/L的NAA时,转基因植株生根率为48.14%-62.13%,对照为14.81%,表明单独使用NAA没有促进不定根发育;添加0.1 mg/L 的IBA时,转基因植株生根率可以达到63.62%-82.95%,对照也提高到29.63%;同时添加0.1 mg/L IBA和0.1 mg/L NAA时,转基因植株生根率可以达到94.75%-100%,生根时间提前15d,对照提高到48.15%,说明在NAA存在时,不定根对IBA的敏感性提高。激素含量测定发现,在添加NAA和IBA时,转基因株系的IAA含量增加,细胞分类素的含量最低,转录组分析的结果表明IAA合成途径中的YUCCA6显著上调表达,信号转导途径中PIN2显著上调表达,而细胞分裂素合成途径中CYP735A1显著下调表达。

图13. CbNN1基因的克隆和载体构建
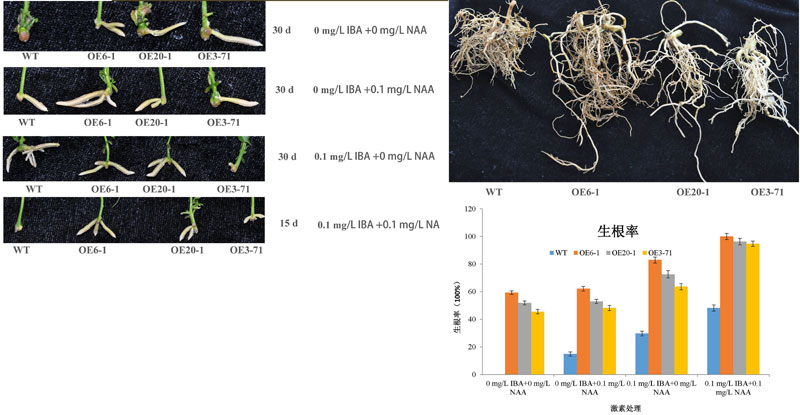
图14. CbNN1基因的表型分析